Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Синтез кетоновых тел
ОБМЕН ЛИПИДОВ (2-ая лекция).
Синтез фосфолипидов (ФЛ) протекает в цитоплазме и на первых стадиях совпадает с синтезом ТАГ. На уровне фосфатидной кислоты и ДАГ эти процессы расходятся. Возможны 2 пути синтеза ФЛ.
1-ый путь характеризуется участием фосфатидной кислоты:
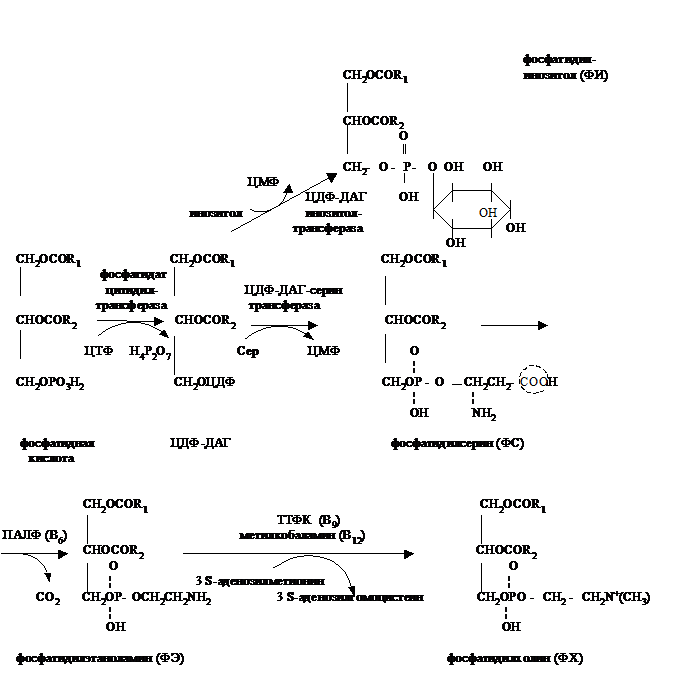
2-ой путь характеризуется участием в синтезе ДАГ и начинается с активации спирта, например холина:

В ФЛ обычно R1 – остаток насыщенной длинноцепочечной жирной кислоты, а R2 – полиненасыщенной, н-р арахидоновой.
Т.к. для синтеза ФЛ и ТАГ нужны общие субстраты, то между этими процессами существует конкуренция. Вещества, способствующие синтезу ФЛ и препятствующие тем самым отложению ТАГ (в тканях, не являющихся жировым депо, например в печени), называются липотропными факторами. К ним относят:
1) структурные компоненты ФЛ – холин, инозитол, серин, этаноламин;
2) ЦТФ, необходимый для образования коферментов – ЦДФ-ДАГ и ЦДФ-холина;
3) В6 (ПАЛФ), нужный для декарбоксилирования ФС с образованием ФЭ;
4) метионин - донор метильных групп для синтеза ФХ из ФЭ;
5) В9 (ТГФК) и В12 (метилкобаламин), участвующие в переносе СН3-групп при образовании ФХ из ФЭ.
Синтез кетоновых тел.
К кетоновым, или ацетоновым, телам относят ацетоацетат, b-гидроксибутират и ацетон. Они являются недоокисленными промежуточными продуктами распада жирных кислот и углеродных скелетов кетогенных аминокислот (Лей, Иле, Лиз, Фен, Тир, Трп), синтезируются из ацетил-КоА. Кетогенез происходит только в МХ печени. Возможны 2 пути синтеза кетоновых тел: гидроксиметилглутаратный цикл (наиболее активный – см. ниже) и деацилазный (учебник).
 |
Образовавшийся ацетил-КоА вступает в 1-ю реакцию - цикл замыкается.
Из ацетоацетата образуются и др. кетоновые тела:
β-гидроксибутират-
дегидрогеназа



 СН3 С СН2 СООН СН3 СН СН2 СООН
СН3 С СН2 СООН СН3 СН СН2 СООН
 О НАДН2 НАД+ ОН
О НАДН2 НАД+ ОН
СО2 β-гидроксибутират

 СН3 С СН3
СН3 С СН3
О
ацетон
Биологическая роль кетоновых тел. Кетоновые тела из печени поступают в кровь (N = 0,1-0,6 ммоль/л), а с ней – во внепеченочные ткани (сердце, легкие, почки, мышцы, нервная ткань), где используются в качестве энергетических субстратов.
Вначале активируется ацетоацетат. Возможны 2 способа:
ацетоацетат-КоА-трансфераза
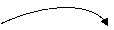

 1) СН3 С СН2 СООН СН3 С СН2 С ~SКоА
1) СН3 С СН2 СООН СН3 С СН2 С ~SКоА
О сукцинил-КоА сукцинат О О
ацетоацетат ацетоацетил-КоА
ацетоацетил-КоА-
синтетаза О

 2) СН3 С СН2 СООН+HSКоА+АТФ СН3 С СН2 С ~SКоА+АМФ+Н4Р2О7
2) СН3 С СН2 СООН+HSКоА+АТФ СН3 С СН2 С ~SКоА+АМФ+Н4Р2О7
О О
Образующийся ацетоацетил-КоА тиолазой (ацетил-КоА-ацил-трансферазой) расщепляется на 2ацетил-КоА:
тиолаза



 СН3 С СН2 С ~SКоА 2 СН3 С ~SКоА ц. Кребса.
СН3 С СН2 С ~SКоА 2 СН3 С ~SКоА ц. Кребса.
О О HSКоА О
Ацетон выводится легкими. b-Гидроксибутират превращается в ацетоацетат:
β-гидроксибутират-
дегидрогеназа


 СН3 СН СН2 СООН СН3 С СН2 СООН
СН3 СН СН2 СООН СН3 С СН2 СООН
ОН НАД+ НАДН2 О
β-гидроксибутират ацетоацетат
Скорость окисления кетоновых тел во внепеченочных клетках пропорциональна их содержанию в крови. Они окисляются активнее по сравнению с глюкозой и СЖК.
Синтез холестерина (ХС)
протекает в цитоплазме многих тканей, наиболее активно – в печени, тонком кишечнике и коже. Синтезируется ХС из ацетил-КоА в 3 этапа:
1) образование мевалоновой кислоты из ацетил-КоА;
2) образование «активного изопрена» и сквалена;
3) превращение сквалена в ХС.
1-ый этап: до образования ГМГ-КоА протекают те же реакции, что и при синтезе кетоновых тел (но вне МХ):
 СН3 О
СН3 О
 ГМГ-КоА-редуктаза
ГМГ-КоА-редуктаза


 ацетил-КоА НООС СН2 С СН2 С ~SКоА
ацетил-КоА НООС СН2 С СН2 С ~SКоА
2 НАДФН2 2 НАДФ+
ОН
 СН3
СН3

 НООС СН2 С СН2 СН2ОН + НSКоА
НООС СН2 С СН2 СН2ОН + НSКоА
ОН
мевалоновая кислота
Реакция необратима, является ключевой, скоростьлимитирующей, активность фермента (1)повышается при дефосфорилировании (инсулин) и снижается при фосфорилировании (глюкагон) и (2)ингибируется холестерином по принципу обратной связи
2-ой и 3-й этапы: мевалоновая кислота с затратой энергии АТФ превращается в «активный изопрен»:
АТФ АТФ

 конденсация циклизация
конденсация циклизация




 мевалоновая кислота «активный изопрен» сквален
мевалоновая кислота «активный изопрен» сквален




 ланостерин ХС
ланостерин ХС
 |  | ||

 3 7
3 7
 ОН
ОН
 С О
С О
R
Гидроксилирование стероидного кольца происходит в монооксигеназной цепи ЭПС.
Часть ХС этерифицируется по 3-ему положению. В клетках, например печени, слизистой кишечника, реакцию катализирует ацил-КоА холестерол-ацилтрансфераза (АХАТ):
АХАТ
 Ацил-КоА + ХС ЭХ + КоА
Ацил-КоА + ХС ЭХ + КоА
В плазме крови ЭХ образуются под действием ЛХАТ (см. «Сл. белки»).
Биологическая роль ХС и его эфиров:
1) построение мембран;
2) из него синтезируются биологически активные вещества стероидной природы:
а) стероидные гормоны (например, гормоны коры надпочечников),
б) витамин Д3 (холекальциферол) – в коже,
в) желчные кислоты – в печени. Их образование включает реакции частичногоокисления боковой цепи и гидроксилирования ХС. Синтез желчных кислот начинается с 7a-гидроксилирования ХС в микросомах, и именно эта реакция является скоростьлимитирующей. Активность 7a-гидроксилазы тормозится по обратной связи желчными кислотами, а повышается при фосфорилировании.
С желчью ХС (как в свободной форме, так и в виде желчных кислот) поступает в кишечник и частично выводится из организма. В кишечнике из первичных желчных кислот образуются вторичные. 98-99% желчных кислот, поступивших в кишечник, возвращаются через систему воротной вены в печень, т.е. наблюдается кишечно-печеночная циркуляция. Потеря с фекалиями 1-2% желчных кислот восполняется синтезом их новых молекул.
Date: 2015-07-17; view: 1424; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |